
- About Us
- Information
-

The Author ensures that the research has been conducted responsibly and ethically with adherence to all relevant regulations. read more..
- For Authors
- For Reviewer
- Manuscript Guidelines
- Membership
- Publication Ethics
-
- Journals
- Reprints
- e-Books
- Videos
- Policies
- Contact Us
 COVID-19
COVID-19
- Submissions



Full Text
Journal of Biotechnology & Bioresearch
Antagonistic Activity of Crude Cellulase (β-glucosidase) Produced by Trichoderma Harzianum Against Some Fungal Pathogens of Oil Palm
Muhammad Muhammad1*, Roswanira Abdul Wahab2, Fahrul Huyop3, Mohd Hefni Rusli4, Naji A. Mahat5, Habeebat Adekilekun Oyewusi6, Umar Aliyu Abdullahi1, Muhammad Ali Dikwa1, Kamisu Shuaibu Idris1, Rabiu Salihu1 and Saminu Ali kofar Kwaru1
1 Department of Microbiology and Biotechnology, Federal University Dutse, Jigawa, Nigeria
2 Department of Chemistry, Faculty of Science, University Technology Malaysia (UT.M)
3 Department of Bioscience, Faculty of Science, University Technology Malaysia (UT.M)
4 Plant Pathology and Bio security Unit, Malaysia Palm Oil Board, Selangor, Malaysia
5 Center for Sustainable Nanomaterials, University Technology Malaysia (UT.M)
6 Department of Biochemistry, School of Science, and Computer Studies, Federal Polytechnic Ado Ekiti, Nigeria
*Corresponding author:Muhammad Muhammad, Department of Microbiology and Biotechnology, Federal University Dutse, Jigawa State, Nigeria
Submission: April 27, 2026;Published: June 05, 2026

Volume6 Issue 2May 06, 2026
Abstract
This study evaluated the antagonistic potential of Trichoderma harzianum Rifai culture and its cellulolytic β-glucosidase against four phytopathogenic fungi of oil palm: Fomitopsis. meliae, Bipolaris sorokiniana, Phoma herbarum, and Ganoderma. boninense. Antifungal activity was assessed through dual culture test, hyperparasitism interactions, and enzymatic degradation of pathogen cell-walls by enzymes produced via solid state fermentation. In dual culture test, T. harzianum Rifai inhibited mycelial growth by 92.80%, 88.70%, 80.20 %, and 98.90% against F. meliae, B. sorokiniana, P. herbarum, and G. boninense respectively, compared to control 99.90% growth in the control. Microscopic analysis of hyperparasitism revealed cell lysis of the target fungi following direct interactions with T. harzianum mycelia, indicating enzymatic digestion of fungal cell walls. Among the cell wall-degrading enzymes secreted, β-glucosidase exhibited the highest activity, with peak value of 199.3U/g, 197.6U/g, 197.8U/g, and 209.5U/g for the respective pathogens. The enzyme significantly suppressed fungal growth on agar plates. The Minimum Inhibitory Concentration (MICs) of T. cellulolytic β-glucosidase 128, 512, 32, and 64mg/mL, while the Minimum Fungicidal Concentrations (MFCs) were 32, 128, 512, and 64mg/mL, respectively. In vivo assays demonstrated that oil palm seeds treated with T. harzianum Rifai cellulolytic β-glucosidase provided superior protection to seedlings compared to soil incorporation methods. Collectively, these findings demonstrate that T. harzianum Rifai-derived cellulolytic β-glucosidase possesses strong antifungal activity and represent a promising biocontrol agent for managing fungal diseases in oil palm..
Keywords:Biocontrol; β-glucosidase; Phytopathogens; Trichoderma harzianum rifai; Cellulolytic; Oil palm; Minimum inhibitory concentration; Minimum fungicidal concentration
Introduction
Fungal diseases affecting oil palm (Elaeis guineensis) plants are the general challenge for various oil palm-producing countries globally, causing serious economically important yield loss of oil palm plants [1-3]. It has cost several Southeast Asian countries as much as USD 500 million in total annual economic losses [4]. According to the review of related literature, the economic loss caused by fungal plant diseases in Indonesia and Malaysia in particular is up to RM 1.5 billion in a year [5]. Oil palm is an oil-producing crop which become the most important commodity plant in Malaysia, with the oil palm industry being the fourth highest contributor to the Malaysian Gross Domestic Product (GDP) [6-8]. Malaysia is also the world’s second-largest oil palm producer, with 5.85 million hectares of oil palm plantation that encompasses over 60% of the agricultural land [9]. Even though oil palm is the major economic plant, the industry faces serious challenges from fungal plant diseases that threaten current plantations. Therefore, specific research on these devastating fungal diseases and their management strategies must be augmented.
Fungi affecting the oil palm plants include Ganoderma boninense, Fusarium sp., Phoma herbarum, Curvularia, Armillaria, Pestalotiopsis, Bipolaris sorokiniana, and Fomitopsis sp. Among these fungi, G. boninense is the major devastating fungal pathogen. This fungal infection in the oil palm leads to upper stem and basal stem rots [10]. Ganoderma is a white-rot fungus from the division Basidiomycetes club fungi; they include common mushrooms [11]. It has been reported as the most disturbing, widespread fungi affecting the oil palm trees mostly occurs in Indonesia and Malaysia; it also occurs in Africa, Papua New Guinea, and Thailand at lesser extent. Ganoderma debilitating plant disease is initiated by the fungi colonizing the oil palm roots and breaking down the basal stem tissues [12-15]. The pathogens then progress and damage the vascular tissues of the oil palm plant, leading to an interrupted water and mineral salts supply from the roots to other parts of the oil plant [16]. Further, necrosis or chlorosis appears on the oil palm, followed by wilting, dried leaves, unopened spears leave, development of fruiting bodies (basidiocarps) on the stem, and eventual death of the tree [17,18]. Moreover, G. boninense infections typically spread through the plants’ root systems [19].
Currently, a variety of disease control strategies to curb fungal infections of oil palm trees that include physical (burning), cultural (soil mounding), regulatory (replanting), and chemical (fungicides) treatments are in place. However, none of these methods has been effective in eradicating and preventing the further proliferation of fungal diseases in oil palm [20]. Current disease management techniques must be combined with other control measures (low doses of pesticides, biological agents) to intensify the killing efficacy. Then again, opting for this more complex technological support system tends to elevate the plantations’ management costs [21]. Another concern is the natural bioaccumulation of pesticides into our food chain and its eventual ingestion by mammals and birds [22,23]. The prevalent and extensive use of chemicals can invariably lead to serious environmental pollution. Worryingly, the indiscriminate use of chemical fungicides and release of large quantities of hazardous compounds into the environment can have deleterious consequences on human health [24], and give rise to more pesticide-resistant microorganisms.
Recent findings highlighted certain fascinating bioactive compounds produced by antagonistic fungus Trichoderma harzianum Rifai inhibited the pathogenic fungus [25]. The fungus was shown to produce a β-glucosidase that hydrolyzed the β-linkage of the amorphic β-1, 3- glucan filling material of the pathogenic fungus chitin-based cell wall [26]. The findings were further proven by an In-silico investigation, where β-glucosidase was shown to facilitate the T. harzianum Rifai mycelia to penetrate the cell wall of fungal pathogen [27]. Driven by this idea, the present study aimed to use the antagonistic T. harzianum Rifai as a greener and sustainable microbial agent to control brown rot disease caused by F. meliae, root rot caused by B. sorokiniana, leaf spot disease and white-rot diseases caused by P. herbarum and basal stem rot caused by G. boninense in oil palm.
Material and Methods
Sampling and collection of fungal isolates
This study utilized previously isolated T. harzianum Rifai reported elsewhere. Three pathogenic fungi (G. boninense, B. sorokiniana and P. herbarum) were collected from the Plant Pathology and Biosecurity Unit of the Malaysian Palm Oil Board (MPOB). The old culture of the fungi strains was sub-cultured on freshly prepared Potato Dextrose Agar (PDA) medium and incubated at 30 ℃ for 7 days. Furthermore, pathogenic fungus (F. meliae) used in this study was isolated from an infected stem, and tissues of a mature oil palm tree as reported by [28]. Each fresh culture was sub-cultured on PDA slants and stored in a refrigerator until use.
In vitro antagonistic activity of T. harzianum Rifai by dual culture and hyperparasitism interactions
In this study, the dual culture interaction study was conducted as described by [29]. The 5mm discs of both fungal pathogens and the antagonist (T. harzianum Rifai) mycelia were cut from the margin of actively growing 7-day old fungi isolates and were placed at 1cm distance from the edge of petri dish containing PDA. The plates were incubated at 28 ℃ for 7 days, and the diameter of the Zone of Inhibition (ZOI) of fungal pathogen’s growth was measured in cm to denote antagonistic activity. The Growth Inhibition Percentage (GIP) was expressed in terms of inhibition percentage (%) of radial growth of the fungal pathogens in relevance to the control plates (5mm disc of the plant pathogens without the presence of T. harzianum Rifai). The growth inhibition percentage (%) was calculated using the formula bellow; as previously described by [30].
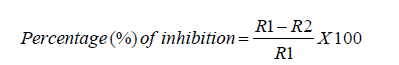
R1 refers to the radius of the radial growth of the pathogen towards the opposite side in the control plate. The term R2 describes the pathogens’ radius of radial growth towards the antagonist in the test plate.
Hyperparasitism interactions were conducted by using the mycelia (100mg) of both fungal pathogens and that of the antagonist cut from the interaction area in dual-culture test plates and fixed for 3h at 4 ℃ in 2% (v/v) glutaraldehyde and 2% (w/v) paraformaldehyde buffered with 0.05% sodium acetate buffer, pH 7, and post-fixed in 1% Karnovsky fixative solution in the same buffer. The mycelia (100mg) were washed with buffer and dehydrated in 25-100% (v/v) ethanol then kept in a desiccator for 24h to dry. Sample preparation for Scanning Electron Microscope (SEM) was conducted as described by [31]. The dried mycelia (100mg) were gold coated in an ion sputter coater and observed at 3000 x magnification under Scanning Electron Microscope (SEM) to visualize enzyme-related disruptions in the mycelia of the fungal pathogens tested.
Production of T. harzianum Rifai crude cellulase and enzyme assay
Crude cellulase was produced using Oil Palm Frond Leaves (OPFL) as suitable substrate via Solid State Fermentation (SSF). In this study, OPFL was grinded into small powdery form of about 2-3mm particle size with help of table grinder (Wellmac RT-08, Taiwan) for the production of cellulase. Production of cellulase was conducted as described by [32,33] with slight modification using 250mL Erlenmeyer flask at 30 ℃, containing 5g of OPFL substrate and 20mL of modified Mendel production media ((NH4)2SO4 1.4g/L, KH2PO4 2.0g/L, urea 0.3g/L, yeast 1.25g/L, CaCl2 0.3g/L, MgSO4.7H2 O 0.3g/L, FeSO4.7H2 O 0.005g/L, MnSO4.H2 O 0.0016g/L, ZnSO4.7H2 O 0.0014g/L, CoCl2 0.002g/L, peptone 1.0g/L and 2mL of Tween 80) pH 5.2. The moisture content of the mixture was read at 80% with the help of moisture analyzer (MX50, A&D Weighing Co., Ltd., Japan). The Erlenmeyer flasks containing the mixture were autoclaved (15min at 121 ℃, 20 psi) and allowed it to cool at 25 ℃. Fermentation of the content begins by inoculation with 2mL of T. harzianum Rifai spore suspension (2.0 x 108 spores/g of OPFL) and kept at 30 ℃ for 7 days under non-optimized conditions. The mixture was collected at 24h intervals and determined the crude extract (cellulase) activity.
In this study, endoglucanase (CmCase) activity was determined by the colorimetric method using carboxymethyl-cellulose as substrate following the method described by [34,35]. One unit of endoglucanase activity is defined as 1μ mole of glucose released per mL of enzyme per minute, under assay conditions. For exoglucanase (Fpase), the activity was determined by colorimetric method using Whatman No.1 filter paper No. 1 strip (1x6cm, 50mg) and kept at 50 ℃ with 1mL sodium acetate buffer (0.05M, pH5) and 0.5mL of crude enzyme cocktail for 60 minutes. Quantification of the released reducing sugars was conducted following the method described by [36]. Then, the absorbance was read at 549nm. One unit of exoglucanase activity was defined as 1 μmole of glucose released per mL of enzyme per minute, under assay condition. For β-glucosidase, the activity was determined also by colorimetric method using PNPG as substrate. The activity was determined based on the released of P-Nitrophenol (PNP) from the PNPG substrate and the absorbance was read at 430nm. One unit of β-glucosidase activity was expressed as the amount of enzyme that released 1μ mole of PNP per min under assay condition. The T. harzianum Rifai grow in modified Mendel production medium without the mycelia of the fungal pathogens were used as control.
Plate inhibition assay of T. harzianum Rifai crude cellulase (β-glucosidase) on pathogenic fungal growth
Plate inhibition assay was performed following a method described by [37,38] with slight modifications, by using agar well technique. In which, the antifungal activity of the crude extract (β-glucosidase) of T. harzianum Rifai against the phytopathogenic fungal growth was conducted in a sterilized petri dish containing 20ml of PDA as the substrate, then 100μl of the supernatant of the fungal pathogens (2.0x104 spores/ml) was spread plated uniformly on the PDA. The agar was punched using sterilized pipette tip to formed wells on the plate at 2cm intervals. The wells were loaded with T. harzianum Rifai β-glucosidase at different concentrations (16, 32, 64, 128, 256, and 512mg/mL). Control wells containing 20μl antifungal agent (imidazole) as positive control and 20μl Dimethyl Sulfoxide (DMSO) in separate wells were used as the negative control. The plates were incubated at 30 ˚C for 72h. The diameters of the halo zones, which indicate complete inhibition on the four tested fungal pathogens (F. meliae, B. sorokiniana, P. herbarum, and, G. boninense) were measured by comparing with the diameter of control. The diameter of the zone of inhibition of the mycelia was calculated using the following formula:
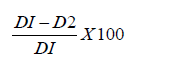
Where DI=diameter of radial growth of fungal pathogen in control plates, D2=diameter of radial growth of the fungal pathogen in treatment plates.
Determination of Minimum Inhibitory Concentration (MIC) and Minimum Fungicidal Concentration (MFC) of T. harzianum Rifai crude cellulase (β-glucosidase) against the fungal pathogens tested
The Minimum Inhibitory and Minimum Fungicidal Concentrations (MIC and MFC) was determined as described by [39- 41] with slight modification, by using a broth macro dilution method. 1mL of different concentrations of T. harzianum Rifai β-glucosidase extract were incorporated into the test tubes containing 10ml of PDB growth media to obtain a concentration of 1000μg/ml stock solution, followed by two-fold-dilutions to obtain (16, 32, 64, 128, 256, and 512mg/mL). 20μl standardized suspension of each of the four tested fungal pathogens (2.0x104spores/ml) was transfer to each test tube. The control test tubes containing 10mL of PDB only without the fungal pathogen, the culture test tubes were incubated at 30 ℃ and read at 595nm when the turbidity of the growth control tube was observed (day-1 reading). After 24h, (day-2 reading), followed by 48h (day- 3 and day 4 readings) and day 5. Minimum inhibitory concentrations expressed in μg/mL were defined as the lowest concentration of the crude T. harzianum Rifai cellulase extract, which did not reveal any growth of the fungal pathogen tested after incubation.
For the Minimum Fungicidal Concentration (MFC), all the tubes that did not reveal any fungal growth after incubation at 30 ℃ from the minimum inhibitory concentration tubes, 100μl of the suspension were taken and plated on freshly prepared PDA, and then the plates were incubated at 30 ℃ for 7 days. The spores of Trichoderma harzianum Rifai were harvested by the addition of 4mL of sterilized distilled water and 1mL of sterilized tween 80 into the plates containing T. harzianum Rifai and scraped the mycelia followed by spore counting. The total number of spores of the fungal pathogens was counted with a hemocytometer using the following formula:
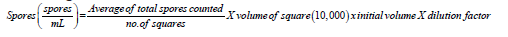
The Minimum Fungicidal Concentration (MFC) was determined as the lowest concentration of the cellulase extract (β-glucosidase) of T. harzianum Rifai that kill (99.5%) mycelial growth of any of the fungal pathogen tested on PDA after incubation period at 30 ℃.
In vivo evaluation of T. harzianum Rifai cellulase (β-glucosidase) efficiency in seeds inoculation and soil incorporation treatments
Efficiency of T. harzianum Rifai cellulase (β-glucosidase) in seeds inoculation and soil incorporation treatments was conducted with some slight changes to previously described methods by [42].
The seeds were coated with slurry (40mL) prepared by mixing 10g of T. harzianum Rifai inoculants with 30mL (40%) gum Arabic (acacia gum) solution (Nexira ®). The inoculant and coated materials were rinsed off the seeds, and the diluent was serially diluted. An aliquot of 0.1mL appropriately diluted T. harzianum Rifai inoculant was spread on PDA plates and incubated at 30 ℃ for 7 days [43]. The artificial inoculation of F. meliae, B. sorokiniana, P. herbarum, and G. boninense fungal pathogens (10mL of 2x104 spores/mL of the fungal pathogens) was performed into the double-autoclaved (121 ℃ 30min) soil mixtures and used as the pathogen-treated in planting trays containing sandy soil, perlite, and peat moss in ratio 1:1:1. Double-autoclaved (121 ℃ 30min) uninoculated soil mixed with compost containing sandy soil, perlite, and peat moss in ratio 1:1:1 was used as the control. The seeds inoculated with fungal pathogens were transferred to a plastic 9cm petri dish containing PDA (Oxoid Ltd, Basingstoke, England) medium. The cell culture plates were then incubated in an incubator for 7 days. The experiment involved 30 treatments performed in triplicate and observed for 6 months.
For the soil incorporation treatment, the T. harzianum Rifai was cultivated in soil containing fermented corn husk and orange peel (1:1; w: w), at a rate of 50g per nursery planting seed trays (300x22x50mm) containing a 1:1:1 ratio of sandy soil, perlite, and peat moss. The soil was sterilized for 1 hour at 121 ℃ and then kept for ten days under greenhouse conditions and watered daily to ensure perfect fermentation. A total of 150g of the mixture was transferred into 1-L jar and autoclaved twice between 24 hours intervals. The suspensions were diluted to 3.5x108 per cm3 conidia concentration before inoculation into the soil. A 50mL/ m2 application rate was used for the nursery planting seed trays (300x22x50mm) in the present study.
In this study, severe chlorotic or necrotic over one-half (<50%) of the leaves, extensive leaf desiccation, and stunted growth of oil palm seedlings were observed. Equation (1) was used to estimate The Plant Disease Index (PDI). The PDI represents the percentage (%) ratio of the total disease rating and production of the sum of plant seedlings and the highest rating value. The diseased area was evaluated up to 50%, while healthy area was represented by more than 50%. The PDI rating scale used in this study was as follows: 0=without any symptoms in oil palm plant; 1=symptoms of about 1-5% in oil palm plant; 2=5-15% symptoms in oil palm plant; 3=15- 50% symptoms in oil palm plant; 4=< 50% symptoms in oil palm plant and the highest rating value is 4<50% symptom in oil palm plant.
Statistical analysis of soil incorporation and seed inoculation treatments
With the help of IBM SPSS version 22.0, the data was statistically analyzed. The normality of the data used for statistical inference for the efficiency of T. harzianum Rifai cellulase (β-glucosidase) in seeds inoculation and soil incorporation treatments were analyzed using the non-parametric test (Mann-Whitney) because the data violated the assumption of normality by considering the significance level of 0.05% using Shapiro-Wilk as described by [44,45]. This was followed by a comparison between the treatments (Treatment 1 seed inoculation and Treatment 2 soil incorporation).
Result and Discussion
In vitro antagonistic activity of T. harzianum Rifai by dual culture and hyperparasitism interactions
In this study, the antagonistic potential of T. harzianum Rifai against the four fungal pathogens tested was examined by dual culture and hyperparasitism interactions (Figure 1). Here, the in vitro dual culture assay results affirmed the efficacy of the T. harzianum Rifai culture to inhibit the growths of the four fungal pathogens tested. Interaction in the dual culture started after two days of incubation. Later, T. harzianum Rifai overgrow the pathogens and sporulated (Figure 1aii,bii,cii&dii) which is contrarily to the growth observed in control (Figure 1ai,bi,ci&di). Additionally, statistical analysis revealed that T. harzianum Rifai exhibited high significance differences with more than 80% inhibition zone (Table 1). The Zone of Inhibition (ZOI) percentage observed was in the following descending order G. boninense > F. meliae, > B. sorokiniana, > P. herbarum having the percentage of growth inhibition 98.90, 92.80, 88.70, and 80.20 % with corresponding median ranges of 2.90-3.97, 2.07-3.97, 2.32-3.97 and 2.17-3.3, respectively. Similarly, not significance 99.90% growth of inhibition with median 3.011 ranges (2.30-3.91) was observed in control (Table 1).
Table 1: Percentage (%) of radial growth of inhibition caused by T. harzianum Rifai fungus in the in vitro dual culture tests against the pathogenic fungi tested.

The percentage of radial growth for the inhibition of four fungal pathogens tested. The data are actual inhibition effects recorded and presented as the median (range). High Significant (HS) percentage of inhibition was observed in all the four pathogens tested. Not Significance (NS) was observed in control inferences at 0.05 level of significance.
Figure 1:Antagonistic ability of T. harzianum Rifai against phytopathogens tested by dual culture and hyperparasitism interactions. Note: (ai) F. meliae control (R1) (aii), F. Meliae + T. harzianum Rifai test (R2), (aiii) F. meliae + T. harzianum Rifai mycelial interaction (bi) B. sorokiniana control (R1) (bii), B. sorokiniana + T. harzianum Rifai tests (R2) (biii) B. sorokiniana + T. harzianum Rifai mycelial interaction (ci) P. herbarum control (R1), (cii) P. herbarum + T. harzianum Rifai test (R2), (ciii) P. herbarum + T. harzianum Rifai mycelial interaction (di) G. boninense control (R1), (dii), G. boninense + T. harzianum Rifai test (R2), (diii) G. boninense+T. harzianum Rifai mycelial interaction. TH represents Trichoderma harzianum Rifai, FM represents Fomitopsis meliae, BS represents Bipolaris sorokiniana, PH represents Phoma herbarum, and GB represents Ganoderma boninense. The yellow color arrow in SEM micrograph 3000x magnification indicated the interaction regions.

This finding corroborated earlier research that suggested several species of the Trichoderma genus were used as biocontrol agents against different pathogenic oil palm’s fungi. The same findings were obtained when T. harzianum Rifai fungus serve as the antagonist against mycoparasite causing charcoal rot in soybean [21]. The antagonistic effect of T. harzianum Rifai fungus seen in this study concurred with the findings on B. sorokiniana reported by [46,47]. Another study on G. boninense causal agent of basal stem rot revealed the effective inhibition caused by T. harzianum sp. reported by [14]. Our findings support the earlier investigation, which found that Trichoderma species had the highest zones of inhibition in dual culture experiments against phytopathogenic fungi as previously reported by [48-51]. Additionally, some strains of Trichoderma sp. were found to be biocontrolling of other pathogenic fungi like F. solani, A. alternata, A. solani, F. graminearum, B. cinerea, F. verticilloides, verticillium, and Sclerotium rolfsii [52- 57]. The outcome of this study indicated that the T. harzianum Rifai fungus appears to be a promising antagonistic agent for inhibiting growths of the four fungal pathogens of the oil palm.
In hyperparasitism interaction, the Scanning Electron Microscopy (SEM) results revealed that the antagonistic behaviors were positively observed from the mycelial interactions of T. harzianum Rifai with that of the four fungal pathogens tested, in which the T. harzianum Rifai mycelia twisted or coiled and penetrated the cell wall of the fungal pathogens as shown in the (Figure 2(a) iii, (b) iii, (c) iii, and (d) iii). This study is consistent with Trichoderma hyphal interactions with fungal pathogens previously reported by [58-61].
It has been reported that, T. harzianum Rifai fungus is a potent producer of cell wall degrading enzymes (CWDEs) [61]. Also, the fungus reportedly liberates other substances (lactones, alcohols, polyketides, piperazine, acetaldehyde, isocyanide, and terpene derivatives) that function as plants growth promoters [21,62,63]. As a matter of fact, this characteristic feature of the T. harzianum Rifai fungus may prove beneficial to the growth and protection of the oil palm tree. The fungus imparts two roles from this study’s point of view, namely, as a bio-fungicide and growth promoter of growing oil palm seedlings. Our observation agreed with the findings reported by [63,64] on endophytic Trichoderma sp. as biocontrol agent against fungal plant pathogenic fungi.
Enzyme activity of crude cellulase produced by T. harzianum Rifai
In this study, T. harzianum Rifai produced Cell Wall Degrading Enzymes (CWDEs) such as endoglucanase (CMCase), exoglucanase (Pfase), and β-glucosidase when grown in modified Mendel production medium in the presence of F. meliae, B. sorokiniana, P. herbarum, and G. boninense fungal pathogens mycelia using oil palm frond leaves substrate. Figure 2ai-dii described the activities of endoglucanase (CMCase), exoglucanase (Fpase), and β-glucosidase Cell-Wall Degrading (CWDEs) secreted by the T. harzianum Rifai in the presence of the mycelia of fungal pathogens tested. In this study, low activity in all the cell-wall degrading enzymes produced was observed in the crude extract after the growth of T. harzianum Rifai without the mycelia of the fungal pathogens (control).
In this study, β-glucosidase was found to have the highest activities among the cell-wall degrading enzymes produced. In F. meliae fungal pathogen mycelia, the highest β-glucosidase activity observed was (199.3U/g) at day 2 of incubation (Figure 2ai&aii) with a very low activities in control 15.2U/g (Figure 2 ai). For the endoglucanase (CMCase), the highest activity obtained was 160U/g in F. meliae at day 3 of incubation. The least endoglucanase (CMCase) activity obtained was 100U/g in F. meliae fungal pathogen mycelia at day 7 of incubation. Whereas, the highest exoglucanase (Fpase) activities obtained in F. meliae mycelia was 105U/g at day 2 of incubation (Figure 2 ai&aii).
The highest activity of β-glucosidase in the mycelia of B. sorokiniana revealed 197.6U/g at day 3 of incubation with the lowest activity 10.5U/g in control (Figure 2bi&bii). In this study, the highest activity of endoglucanase (CMCase), and exoglucanase (Fpase) was 165 and 130U/g respectively in B. sorokiniana fungal pathogen mycelia observed at day 3 (Figure 2bi&bii). In the mycelia of P. herbarum fungal pathogen, the highest β-glucosidase 197.8U/g at day 2, CMCase 156.7U/g, and Fpase activity 107.7 at day 4 respectively was observed (Figure 2ci&cii). For the G. boninense fungal pathogen, the highest β-glucosidase activities observed in the mycelia was 209.5U/g at day 3, 180.5U/g CMCase activities at day 3, and 134.6U/g activities at day 2 for Fpase activity (Figure 2di&dii). The lowest activity 7.5U/g in control at day 3 was observed (Figure 2di&dii). This proved the influence by the type of fungal pathogen mycelia in the secretion of cell-wall degrading enzymes, and this could be attributed to the various compositions of the fungal pathogen’s mycelia. This study is consistent with findings on the activities of endoglucanase (CMCase), exoglucanase (Fpase), and β-glucosidase produced by Trichoderma sp reported by [32], Also, corresponded to the findings reported by [29] on the enzyme activities produced by T. harzianum sp in a purified cell-wall from fungal pathogens Rhizoctonia solani, Macrophomina phaseolina.
Figure 2:The activities of endoglucanase (CMCase), exoglucanase (Fpase), and β-glucosidase Cell-Wall Degrading (CWDEs) secreted by the T. harzianum Rifai in the presence of fungal pathogens mycelia.

This study proved the influence by the type of fungal pathogen mycelia in the secretion of cell-wall degrading enzymes, and this could be attributed to the various compositions of the fungal pathogen’s mycelia. This study is corroborated with findings on the activities of cellulase reported by [65]. Also, corresponded to the findings on the cellulase activity produced by T. harzianum sp in a plant pathogenic Phytium sp reported by [66].
Cellulase are vital hydrolytic enzymes produced by microbes including fungi, that are responsible for the conversion of cellulose into simple sugars [67-70]. In this study, T. harzianum Rifai was used for the determination of cellulase activity in the mycelia of fungal pathogens tested. It is well-known that Trichoderma sp. is the most potential for cellulase production which is very effective for degrading the cell-wall of the available substrate [61,71]. Majority of pathogenic fungi possess cell-walls consisting of complex polymers of β-1,3 and β-1,6-glucans, where chitin used as structural backbone and glucan serve as amorphous filing material [72,73]. Therefore, degradation of fungal cell-wall needs various hydrolytic enzymes [73]. The presence study used cellulase from T. harzianum Rifai in the degradation of fungal pathogens mycelia. Figure 2ai&aii described the activity of crude extract (cellulase) secreted by the T. harzianum Rifai in the presence of the mycelia of fungal pathogens tested at 7 days of incubation.
Cellulase is potent in the degrading of β-glucans by supporting β-glucanase in the antifungal activity. So, the level of expression of antifungal activity is also attributed to the components of the mycelial cell-wall of the fungal pathogen [51]. Also, this shows the great effect of T. harzianum Rifai crude cellulase in the hydrolysis of fungal pathogen’s mycelia tested which is consisted with findings reported by [74].
Determination of Minimum Inhibitory Concentration (MIC) and Minimum Fungicidal Concentration (MFC) of T. harzianum Rifai crude cellulase (β-glucosidase) against the fungal pathogens tested
The study revealed that F. meliae (50%) and P. herbarum (34%) showed high percentage inhibition zone at 16mg/mL concentration of the extract while B. sorokiniana (44%) and G. boninense (52%) presented high inhibitory potential at 32mg/mL and 512mg/mL, respectively as shown in Table 2. The cellulolytic β-glucosidase of T. harzianum Rifai diffuses into the agar and inhibits the growth of fungal pathogens. This study corresponded to findings on antimicrobial activity of glycosidase inhibitory protein isolated from Cyphomandra betacea send t. fruit reported by [38]. Also, is consistent to findings on Trichoderma virens Gl006 and Bacillus velezensis Bs 006 a compatible interaction controlling Fusarium oxysporum causing vascular wilt in cape gooseberry (physalis peruviana) reported by [37].
Figure 3:Plate inhibition assay showing the percentage (%) of radial growth of fungal pathogens inhibited by the different concentrations of T. harzianum Rifai crude cellulase (β-glucosidase) against a) F. meliae (b) B. sorokiniana (c) P. herbarum (d) G. boninense

Table 2: Percentage of inhibition caused by various concentrations of T. harzianum Rifai cellulolytic β-glucosidase on pathogenic fungi tested.

In this study, the results of the plate inhibition assay using the crude cellulolytic β-glucosidase extract of T. harzianum Rifai showed a positive antagonistic behavior and inhibitory activity at various concentrations (Figure 3). The larger halo zone seen on the enzyme-treated with fungi pathogens could be ascribed to the higher-level secretions of ligninolytic enzymes (Figure 3a-d). Moreover, this could be as a result the release of many microbial metabolites that led to the inhibition and this corresponded with compatible interaction controlling the Fusarium wilt of cape gooseberry reported by Izquierdo-García et al. [37]. The study revealed that cellulase extract (β-glucosidase) of the T harzianum Rifai has decreased the conidial exposure of all the fungal pathogens tested on the plate more than that of control (Figure 3a-d).
In this study, the minimum inhibitory concentration of T. harzianum Rifai cellulase (β-glucosidase) in the inhibition of F. meliae, B. sorokiniana, P. herbarum and G. boninense was determined at the concentration of 128mg/mL in F. meliae, 512mg/mL in B. sorokiniana, 32mg/mL in P. herbarum and 64mg/mL in G. boninense (Figure 4) with high activity 82, 76, 70, and 84 U/mL in F. meliae, B. sorokiniana, P. herbarum, and G. boninense respectively compared to control with high enzymatic activity 45U/mL (Figure 4). The activity of different concentrations of T. harzianum Rifai cellulase (β-glucosidase) for the inhibition of fungal pathogens tested in the present study was depicted in Figure 4.
Figure 4:Plate inhibition assay showing the percentage (%) of radial growth of fungal pathogens inhibited by the different concentrations of T. harzianum Rifai crude cellulase (β-glucosidase) against a) F. meliae (b) B. sorokiniana (c) P. herbarum (d) G. boninense

Table 3: Fungal spores counted (spores/mL) after incubation period for determining the Minimum Fungicidal Concentration (MFC) in the inhibition of each fungal pathogen tested

The Minimum Fungicidal Concentrations (MFC) of T. harzianum Rifai extract cellulase (β-glucosidase) was determined from the plates containing the less concentration (MIC plates) which are 32mg/mL, 128mg/mL, 512mg/mL, and 64mg/mL observed against F. meliae, B. sorokiniana, P. herbarum and G. boninense respectively. The MFC in F. meliae was observed on day 2 at the concentration of 32mg/mL with the 420,000 spores/mL (Table 3). For the B. sorokiniana, the MFC was observed on day 3 at 128mg/ mL concentration with the 118,000 spores/mL. In P. herbarum, the Minimum Fungicidal Concentration (MFC) was observed on day 4 at the concentration of 512mg/mL with 300,000 spores /mL (Table 3). The MFC in G. boninense was observed at day 5 with 632,000 spores/mL (Table 3).
High activity 3.29, 2.22, 2.01, and 4.60U/mL was observed in F. meliae, B. sorokiniana, P. herbarum, and G. boninense respectively, in comparison to control with activity1.2U/mL (Figure 5).
Figure 5:Minimum Fungicidal Concentration (MFC) for the crude cellulase of T. harzianum Rifai cellulase (β-glucosidase) in the inhibition of fungal pathogens tested and its activity.

In vivo evaluation of T. harzianum Rifai cellulase (β-glucosidase) efficiency in seeds inoculation and soil incorporation treatments
Efficiency of T. harzianum Rifai cellulase (β-glucosidase) in seeds inoculation and soil incorporation treatments was conducted based on the favorable outcome of the in vitro work, the study tested the efficacy of the T. harzianum Rifai cellulase (β-glucosidase) to curb infestations of the F. meliae, B. sorokiniana, P. herbarum and G. boninense pathogens on oil palm seeds and growing seedlings. The test samples’ plant disease index was monitored for growth parameters (shoot length and root length) and seed weight as disease severity indicators. A low plant disease index is desired in this investigation as it represents the low presence or non-existent disease symptoms on the test plants, hence a healthier test subject.
In this study, the oil palm seed were treated with T. harzianum Rifai cellulase (β-glucosidase) then grown in soil inoculated with G. boninense, F. meliae, B. sorokiniana and P. herbarum pathogens. This was based on a recent report which claimed that seed inoculation with bio-fungicide is more effective than the soil incorporation practice in the biological control of oil palm diseases [15]. This is probably due to the longer time required for the T. harzianum Rifai to grow and mature before the fungus could exert substantial antagonistic activity [21]. The study demonstrated that the T. harzianum Rifai inhibited the growth of F. meliae, B. sorokiniana, P. herbarum, and G. boninense in the oil palm seeds due to the present of defense-related enzyme it contained. Such a conclusion was drawn following the assessment of shoot, root, and oil palm seeds between the control and oil palm seedling in the planting tray (treatments). After treatment, the disease symptoms on the negative control (untreated with the crude cellulase) oil palm plant seedlings were noticeable by naked-eye inspection. The root systems of the infected seedlings were damaged (Figure 6B) compared to the negative control (Figure 6A) in which the former was likely due to spongy growth or cellular disruptions caused by the F. meliae, B. sorokiniana, P. herbarum and G. boninense pathogens. The study discovered that shoots and roots of oil palm seedlings treated with the cellulase (β-glucosidase), were markedly longer (5cm-20cm length in shoot; 4cm-10cm length in root and 0.06-0.09g in seed weight) (Figure 6Ai&Bi) than the negative control (shoots length 4-8cm; 2-5cm root length and seed weight 0.03-0.05 g) Figure 6Aii&Bii. This study is consistent with findings on evaluation of Trichoderma isolates as potential biological control agent against soybean charcoal rot disease caused by Macrophomina phaseolina reported by [21].
The present study affirmed that the T. harzianum Rifai cellulolytic β-glucosidase were potent in suppressing growths F. meliae, B. sorokiniana, P. herbarum and G. boninense. Healthier roots and shoots of the seedlings were observed on the cellulase-treated samples (Figure 6Ai&6Bi) compared to the negative control (Figure 6Aii&6Bii), in the planting tray seed inoculation experiment. The plant disease index of the (β-glucosidase) cellulase-treated oil palm roots and seeds (Figure 6Bi&6Cii were substantially different compared to the negative control (Figure 6Aii,Bii&6Ci). The germinated oil palm seeds in this study were depicted in Figure 6D. The above results conveyed that the T. harzianum Rifai enzymatic treatment on the oil palm seedlings protected them from the fungal pathogens based on the defense-related proteins such as β-1,3- glucanase and chitinase that digested the four pathogenic fungal cell walls [75].
Figure 6:Efficacy of T. harzianum Rifai cellulolytic β-glucosidase after treatment in a field. Note: A: Oil palm shoot (Ai: T. harzianum Rifai cellulase (β-glucosidase) seed treatment); Aii: untreated); B: oil palm root (Bi: T. harzianum Rifai cellulase (β-glucosidase) seed treatment; Bii untreated); C: oil palm seeds (Ci: untreated; Cii: T. harzianum Rifai cellulase (β-glucosidase) seed treated; D: germinated seeds.

The hydrolytic action of the cellulase (β-glucosidase) enzyme on the pathogens imparts growth stress on the pathogenic fungi, which inhibited their growth and germination process. This, in turn, increased the likelihood of the oilseeds to grow well. The study concluded that the T. harzianum Rifai cellulase (β-glucosidase) is effective in suppressing the growth of the four fungal pathogens. The cellulase (β-glucosidase) of T. harzianum Rifai efficacy to inhibit the four fungal pathogens in the seed treatment experiment occurred in the following descending order: G. boninense > F. meliae pathogen, > B. sorokiniana > P. herbarum.
Statistical analysis of soil incorporation and seed inoculation treatments
Sequel to the abnormality of the data, the Mann Whitney test of statistic was conducted for comparison between treatment seed inoculation and soil incorporation treatments. The data were analyzed, and the effects of cellulase (β-glucosidase) of T. harzianum Rifai were determined in all the two treatments when compared with that of controls as well as between treatment 1(seed inoculation) and treatment 2 (soil incorporation). In the present study, the effect of cellulase (β-glucosidase) of T. harzianum Rifai were separately recorded for both seed inoculation and soil incorporation treatments, followed by comparison between treatments and the data of the effects and that of controls were recorded. The results of statistical analysis between the treatments (seed inoculation and soil incorporation) are shown in (Table 4).
Table 4: The efficacy of Trichoderma harzianum Rifai cellulase (β-glucosidase) for the seed inoculation and soil incorporation studies.

The effects of T. harzianum Rifai cellulase (β-glucosidase) on fungal pathogens (F. meliae, B. sorokiniana, P. herbarum, and G. boninense) of oil palm seedlings. The data of the actual effects are recorded and presented as median (range). Mann-Whitney U test was used for comparing the parameters between control and treatments (either seed inoculation or soil incorporation), as well as between treatments with the level of significance of 0.05. HS and S represent highly (p<0.001) and significant differences (p<0.05) for comparisons between controls and individual treatment, respectively. In addition, HS* represents the highly significant difference (p<0.001) between the two treatments.
In this study, high significance (<0.001) was observed in shoot length treated with seed inoculation (treatment 1) having the median 8.30, range (2.0-3.0) and significant in seed weight was also observed in seed inoculation treatment with median 1.50, range (0.3-4.0) (Table 4). In treatment 2 (soil incorporation) the growth rate of the shoot length was also significant (<0.001) having the median 10. 00, range (0.1-43.7) (Table 4). Another high significance (<0.001) in the growth rate of root length with the median 10.01 range (5.5-47.6) was observed after soil incorporation treatment (Table 4). In this study, the seed weight having the median 1.39, ranges (0.3-10.3) after treatment with soil incorporation revealed a high significance (<0.001). Whereas the Plant Disease Index (PDI) in both treatments (seed inoculation and soil incorporation) revealed a high significance difference <0.001 at 0.05% significant level (Table 4), in which the percentage of PDI observed in seed inoculation was 9.63, range (1.0-14.0), while in soil incorporation (treatment 2), the PDI percentage observed was 19.65, with different range (2.5-20.0) (Table 4).
In comparison to the results of soil incorporation with that of the inoculation of the seeds, the seeds inoculation approach with large variance (61.91) in shoot length and low plant disease index 9.63% in treatment 1 (seed inoculation) produced better results than the soil incorporation having the variance in shoot length (54.84) and plant disease index 19.65%, compared to control 43.21 and 33.14 PDI% in seed and soil treatments respectively. The root length revealed highly significant growth rate among all the growth parameters tested in the present study (Table 4). The favorable outcome seen here supports the protective effect of the T. harzianum Rifai cellulase (β-glucosidase) on the seeds, as seen in the significant increase in root length (p-value<0.001). The results clearly indicated the superiority of the seed inoculation treatment over the soil incorporation.
Seed inoculation in plant protection has been reported by various researchers Gmell et al. (2005) described that the various seeds pellet pH, seeds toxicity, seeds nodulation, and rhizssobial counts could be evaluated from the grow-out test. The minimum rhizobial number for this test was 8400 per seed, for untreated lucerne was 1380 per seed, and >100 per seed for the subterranean clover, white and red clovers alongside other miscellaneous species. Seeds with a good growth rate were observed in the uninoculated seed of as much as (73%), with minor subterranean clover of about (32%) and less white clover (3%), red clover (4%) with growth in miscellaneous species (0%). Similarly, in the seed inoculation method investigated by this study, the enzyme-treated oil palm seedlings in the planting tray experiment revealed a higher increase in shoot length (5.00-20cm). Significant effect was observed in both soil incorporation and seed inoculation methods as well as in control. Hence, the findings strongly support the efficacy of the enzyme treatment in controlling fungal plant diseases caused by F. meliae, B. sorokiniana, P. herbarum, and G. boninense especially in oil palm plants.
The outcomes obtained from both in vitro and planting tray studies showed that T. harzianum Rifai might be a promising biofungicide to inhibit F. meliae, B. sorokiniana, P. herbarum, and G. boninense infestations in oil palm seeds and seedlings. In this study, the growth factors such as seeds weight, shoot-and root lengths were used to measure the growth rates of the oil palm seedlings and the intensity, while the technique of PDI rating scale evaluated the disease severity. Increase in seeds weight alongside shoot- and root lengths of the seedlings were observed in this study.
The efficacy of cellulase (β-glucosidase) of T. harzianum Rifai in the seed inoculation- and soil incorporation treatments on fungal pathogens viz; F. meliae, B. sorokiniana, P. herbarum and G. boninense fungal pathogens of oil palm plant appear appropriate. As efficacies of the (β-glucosidase) cellulase-treated samples (seed inoculation and soil incorporation) were well-fitted within the previously prescribed acceptable significance level (5%) [21,76].
The implicit impact of the cellulase (β-glucosidase) treatment for oil palm seeds and seedlings protection involved the acceleration of oil palm seeds and seedlings vigor, increase in seed and seedlings aging, increase of radicle and hypocotyl length, and influence the rate of seeds germination. In addition to that, seeds treated with crude cellulase of T. harzianum Rifai enhances the growth of oil palm seedlings and increase the seed’s size and decrease the oil palm seed dormancy. Seeds treated with enzymes, in our own case cellulase (β-glucosidase) of T. harzianum Rifai increase the damaged cells of the oil palm seeds and enhances seed quality. Similarly, the oil palm seeds treated with extracellular enzyme such as cellulase accelerate the degradation of protein network and support the conversion of the complex seed lipoprotein molecules into simple lipid and protein molecules then enhance the seed quality. The potential ability of seed born fungal pathogens in oil palm seed depends heavily on the integrity of cell walls degrading enzymes enhances the cell structure, and increase the seed quality for maximum protection of oil palm seeds as well as to give successful oil palm seedlings growth.
This revealed a significant effect in both seed and soil incorporation compared to control. In which the inoculation treatment was better in protecting the seeds and their growth in seedlings compared to the soil incorporation treatment. This was probably because the oil palm seeds are smaller in size, hence the lower surface coverage of the cellulase enzyme to give protection. Conversely, the seedlings are larger, and the enzyme treatment may not be homogeneously applied throughout the whole plant. The study noted that the applied cellulolytic β-glucosidase permeated through the fungal pathogens’ seed and soil suppressed growths. Since the study also drenched the soil with the spore suspension of T. harzianum Rifai, the germinated spores would have imparted long-term protection on the growing seedlings against the tested fungal phytopathogens which is consistent with findings reported by [21].
Conversely, the deformed seedling roots and shoots system seen here was due to active infections by the pathogenic fungi, which disrupted vascular tissues, i.e., xylem and phloem. This interrupted the systemic flow of mineral salts throughout the oil palm seedlings [18], which explained the desiccation, wilting, and the poor growing oil palm seeding. Therefore, its application as an eco-friendly alternative means in controlling diseases due to fungal infestations in oil palm is recommendable. Hence, T. harzianum Rifai is effective when used in seed treatment or soil incorporation in an agricultural setting.
Conclusion
This study proved T. harzianum Rifai and its cellulolytic β-glucosidase showed strong antagonistic activity against the main fungal pathogens of oil palm. T. harzianum revealed a high antifungal activity in vitro, because it inhibited 80-99% of mycelial growth for Fomitopsis meliae, Bipolaris sorokiniana, Phoma herbarum, and Ganoderma boninense in dual culture tests. The hyperparasitism interactions showed direct mycelial attack and cell lysis of the pathogens. In this study, cellulolytic β-glucosidase is key enzyme among the cell-wall-degrading enzymes produced that had the highest activity. It suppressed fungal growth on agar plates, with MIC values of 32-512mg/mL and MFC values of 32-512mg/ mL depending on the pathogen. For the effective in vivo protection, oil palm seeds treated with the cellulolytic β-glucosidase gave better seedling protection than soil in co-operation methods. This suggests practical application at the nursery stage. Therefore, T. harzianum Rifai and its cellulolytic β-glucosidase can serve as a potent, eco-friendly biocontrol agent. Using them could lower the environmental health hazards linked to chemical fungicides. Because they were effective in the laboratory and seedling tests, they can be developed into “novel and safe biofungicides”. The point is that using these biological agents could reduce reliance on synthetic chemical fungicides. Pertinently, the T. harzianum Rifai cellulolytic β-glucosidase offers a safer and environmental-friendly means of in situ management of oil palm fungal phytopathogens.
References
- Korbecka-Glinka G, Piekarska K, Wiśniewska-Wrona M (2022) The use of carbohydrate biopolymers in plant protection against pathogenic fungi. Polymers 14(14): 2854.
- Paterson RR (2019) Ganoderma boninense disease of oil palm to significantly reduce production after 2050 in Sumatra if projected climate change occurs. Microorganisms 7(1): 24.
- Qaim M, Sibhatu KT, Siregar H, Grass I (2020) Environmental, economic, and social consequences of the oil palm boom. Annual Review of Resource Economics 12 (1): 321-344.
- Hushiarian R, Yusof NA, Dutse SW (2013) Detection and control of Ganoderma boninense: Strategies and perspectives. Springer Plus 2(1): 555.
- Ommelna BG, Jennifer AN, Chong KP (2012) The potential of chitosan in suppressing Ganoderma boninense infection in oil-palm seedlings. Journal of Sustainability Science and Management 7(2): 186-192.
- Cui J, Lamade E, Tcherkez G (2020) Seed germination in oil palm (Elaeis guineensis): A review of metabolic pathways and control mechanisms. International Journal of Molecular Sciences 21(12): 4227.
- Onoja E, Chandren S, Abdul RFI, Mahat NA, Wahab RA (2019) Oil palm (Elaeis guineensis) biomass in Malaysia: The present and future prospects. Waste and Biomass Valorization 10(8): 2099-2117.
- Romes NB, Hamid MA, Hashim SE, Wahab RA (2019) Statistical modelling of ultrasonic-aided extraction of Elaeis guineensis leaves for better-quality yield and total phenolic content. Indonesian Journal of Chemistry 19(3): 811-826.
- Maluin FN, Hussein MZ, Idris AS (2020) An overview of the oil palm industry: Challenges and some emerging opportunities for nanotechnology development. Agronomy 10(3): 356.
- Ramzi AB, Me ML, Ruslan US, Baharum SN, Muhammad NA (2019) Insight into plant cell wall degradation and pathogenesis of Ganoderma boninense via comparative genome analysis. Peer J 7: e8065.
- Susanto A, Sudharto PS, Purba RY (2005) Enhancing biological control of basal stem rot disease (Ganoderma boninense) in oil palm plantations. Mycopathologia 159(1): 153-157.
- Dhillon B, Hamelin RC, Rollins JA (2021) Transcriptional profile of oil palm pathogen, Ganoderma boninense, reveals activation of lignin degradation machinery and possible evasion of host immune response. BMC Genomics 22(1): 326.
- Fook-Hwa L, Omar AR, Seman IA, As'wad AWM, Ganesan V, et al. (2021) Enhanced Polyethylene Glycol (PEG)-mediated protoplast transformation system for the phytopathogenic fungus, Ganoderma boninense. Folia Microbiologica 66(4): 677-688.
- Siddiquee S, Yusuf UK, Hossain K, Jahan S (2009) In vitro studies on the potential Trichoderma harzianum for antagonistic properties against Ganoderma boninense. Journal of Food, Agriculture and Environment 7(3&4): 970-976.
- Siddiqui Y, Surendran A, Paterson RR, Ali A, Ahmad K (2021) Current strategies and perspectives in detection and control of basal stem rot of oil palm. Saudi Journal of Biological Sciences 28(5): 2840-2849.
- Sahebi M, Hanafi MM, Wong MY, Idris AS, Azizi P, et al. (2015) Towards immunity of oil palm against Ganoderma fungus infection. Acta Physiologiae Plantarum 37(10): 195.
- Naher L, Yusuf UK, Ismail A, Tan SG, Mondal MM (2013) Ecological status of 'Ganoderma' and basal stem rot disease of oil palms ('Elaeis guineensis' Jacq.). Australian Journal of Crop Science 7(11): 1723-1727.
- Rebitanim NA, Hanafi MM, Idris AS, Abdullah SN, Mohidin H, et al. (2020) GanoCare® improves oil palm growth and resistance against Ganoderma basal stem rot disease in nursery and field trials. BioMed Research International 2020: 3063710.
- Noor AAN, Bejo SK, Jahari M, Muharam FM, Yule I, et al. (2020) Early detection of Ganoderma boninense in oil palm seedlings using support vector machines. Remote Sensing 12(23): 3920.
- Bharudin I, Wahab AAF, Samad MAA, Xin YN, Zairun MA, et al. (2022) Review update on the life cycle, plant-microbe interaction, genomics, detection and control strategies of the oil palm pathogen Ganoderma boninense. Biology 11(2): 251.
- Khalili E, Javed MA, Huyop F, Rayatpanah S, Jamshidi S, et al. (2016) Evaluation of Trichoderma isolates as potential biological control agent against soybean charcoal rot disease caused by Macrophomina phaseolina. Biotechnology & Biotechnological Equipment 30(3): 479-488.
- Edbeib MF, Wahab RA, Kaya Y, Huyop F (2017) In silico characterization of a novel dehalogenase (DehHX) from the halophile Pseudomonas halophila HX isolated from Tuz Gölü Lake, Turkey: Insights into a hypersaline-adapted dehalogenase. Annals of Microbiology 67(5): 371-382.
- Khalili E, Javed MA, Huyop F, Wahab RA (2019) Efficacy and cost study of green fungicide formulated from crude beta-glucosidase. International Journal of Environmental Science and Technology 16(8): 4503-4518.
- Tsatsakis AM, Nawaz MA, Kouretas D, Balias G, Savolainen K, et al. (2017) Environmental impacts of genetically modified plants: A review. Environmental Research 156: 818-833.
- Khalili E, Huyop F, Javed MA, Mahat NA, Batumalaie K, et al. (2018) Assessments on the catalytic and kinetic properties of Beta-glucosidase isolated from a highly efficient antagonistic fungus Trichoderma harzianum. Biosci J 34(4): 830-847.
- Khalili E, Huyop F, Myra AMF, Wahab RA (2017) Optimization of cultivation conditions in banana wastes for production of extracellular β-glucosidase by Trichoderma harzianum Rifai efficient for in vitro inhibition of Macrophomina phaseolina. Biotechnology & Biotechnological Equipment 31(5): 921-934.
- Bahaman AH, Abdul WR, Hamid AA, Halim KB, Kaya Y, et al. (2020) Substrate docking and molecular dynamic simulation for prediction of fungal enzymes from Trichoderma species-assisted extraction of nanocellulose from oil palm leaves. Journal of Biomolecular Structure and Dynamics 38(14): 4246-4258.
- Muhammad M, Wahab RA, Rusli MH, Huyop F (2022) Occurrence and identification of Basidiomycetous Fomitopsis species the causal agent of Brown-rot in oil palm Elaeis guineensis in Johor, Malaysia. Journal of Tropical Life Science 12(1): 21-35.
- Monteiro VN, Nascimento SRD, Steindorff AS, Costa FT, Noronha EF, et al. (2010) New insights in Trichoderma harzianum antagonism of fungal plant pathogens by secreted protein analysis. Current Microbiology 61(4): 298-305.
- Skidmore AM, Dickinson CH (1976) Colony interactions and hyphal interference between Septoria nodorum and phylloplane fungi. Transactions of the British Mycological Society 66(1): 57-64.
- Ezeilo UR, Wahab RA, Tin LC, Zakaria II, Huyop F, et al. (2020) Fungal-assisted valorization of raw oil palm leaves for production of cellulase and xylanase in solid state fermentation media. Waste and Biomass Valorization 11(7): 3133-3149.
- Ezeilo UR, Lee CT, Huyop F, Zakaria II, Wahab RA (2019) Raw oil palm frond leaves as cost-effective substrate for cellulase and xylanase productions by Trichoderma asperellum UC1 under solid-state fermentation. Journal of Environmental Management 243: 206-217.
- Priyanka SB (2019) Isolation, purification and characterization of pectinase enzyme from Streptomyces thermocarboxydus. Journal of Biotechnology & Bioresearch 5(1): 1-6.
- Miller GL, Blum R, Glennon WE, Burton AL (1960) Measurement of carboxymethylcellulase activity. Analytical Biochemistry 1(2): 127-132.
- Sahrawat R, Garg AP (2023) Isolation of cellulolytic microfungi from rice straw and evaluation of their enzymatic degradation potential under Solid-State-Fermentation conditions. J Biotech Biores 4 (4): 572-579.
- Miller GL (1959) Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry 31(3): 426-428.
- Izquierdo-García LF, Moreno-Velandia CA (2024) Modes of action of Trichoderma virens Gl006 and Bacillus velezensis Bs006: Decoding the arsenal of the microbial consortium. European Journal of Plant Pathology 170(1): 39-65.
- Ordóñez RM, Ordóñez AA, Sayago JE, Moreno MI, Isla MI (2006) Antimicrobial activity of glycosidase inhibitory protein isolated from Cyphomandra betacea fruit. Peptides 27(6): 1187-1191.
- Chandrasekaran M, Venkatesalu V (2004) Antibacterial and antifungal activity of Syzygium jambolanum seeds. Journal of Ethnopharmacology 91(1): 105-108.
- Pujol I, Guarro J, Llop C, Soler L, Fernández-Ballart J (1996) Comparison study of broth macrodilution and microdilution antifungal susceptibility tests for the filamentous fungi. Antimicrobial Agents and Chemotherapy 40(9): 2106-2110.
- Pujol I, Aguilar C, Fernandez-Ballart J, Guarro J (2000) Comparison of the minimum fungicidal concentration of amphotericin B determined in filamentous fungi by macrodilution and microdilution methods. Medical Mycology 38(1): 23-26.
- Rusli MH, Idris AS, Cooper RM (2016) Evaluation of Malaysian soils for potential suppressiveness of fusarium wilt of oil palm caused by Fusarium oxysporum f. sp. elaeidis. Journal of Microbial & Biochemical Technology 8(6): 459-464.
- Gemell LG, Hartley EJ, Herridge DF (2005) Point-of-sale evaluation of preinoculated and custom-inoculated pasture legume seed. Australian Journal of Experimental Agriculture 45(3): 161-169.
- Mahat NA, Zainol-Abidin NL, Nordin NH, Abdul-Wahab R, Jayaprakash PT (2016) Patterns of oviposition and development of Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae) and Chrysomya rufifacies (Macquart) (Diptera: Calliphoridae) on burned rabbit carcasses. Forensic Science International 260: 9-13.
- Mahat NA, Yin CL, Jayaprakash PT (2014) Influence of Paraquat on Chrysomya megacephala (Fabricius) (Diptera: Calliphoridae) infesting minced‐beef substrates in Kelantan, Malaysia. Journal of Forensic Sciences 59(2): 529-532.
- Villa-Rodriguez E, Lugo-Enríquez C, Ferguson S, Parra-Cota FI, Cira-Chávez LA, et al. (2022) Trichoderma harzianum sensu lato TSM39: A wheat microbiome fungus that mitigates spot blotch disease of wheat (Triticum turgidum subsp. durum) caused by Bipolaris sorokiniana. Biological Control 175: 105055.
- Yasmin S, Sultana S, Adhikary SK, Jahan N, Rahman S, et al. (2014) In vitro evaluation of Trichoderma harzianum (Rifai.) against some soil and seed borne fungi of economic importance. J Agric Vet 7(7): 33-37.
- Chérif M, Benhamou N (1990) Cytochemical aspects of chitin breakdown during the parasitic action of a Trichoderma on Fusarium oxysporum f. sp. radicis-lycopersici. Phytopathology 80(12): 1406-1414.
- Das MM, Haridas M, Sabu A (2019) Biological control of black pepper and ginger pathogens, Fusarium oxysporum, Rhizoctonia solani and Phytophthora capsici, using Trichoderma Biocatalysis and Agricultural Biotechnology 17: 177-183.
- Hermosa MR, Grondona I, Iturriaga ET, Diaz-Minguez JM, Castro C, et al. (2000) Molecular characterization and identification of biocontrol isolates of Trichoderma Applied and Environmental Microbiology 66(5): 1890-1098.
- Mucha J, Dahm H, Strzelczyk E, Werner A (2006) Synthesis of enzymes connected with mycoparasitism by ectomycorrhizal fungi. Archives of Microbiology 185(1): 69-77.
- Sempere F, Santamarina MP (2009) Antagonistic interactions between fungal rice pathogen Fusarium verticillioides (Sacc.) Nirenberg and Trichoderma harzianum Annals of Microbiology 59(2): 259-266.
- Foroutan A (2013) Evaluation of Trichoderma isolates for biological control of wheat Fusarium foot and root rot. Romanian Agricultural Research 30: 335-342.
- Chowdappa P, Kumar SM, Lakshmi MJ, Upreti KK (2013) Growth stimulation and induction of systemic resistance in tomato against early and late blight by Bacillus subtilis OTPB1 or Trichoderma harzianum Biological Control 65(1): 109-117.
- El-Debaiky SA (2017) Antagonistic studies and hyphal interactions of the new antagonist Aspergillus piperis against some phytopathogenic fungi in vitro in comparison with Trichoderma harzianum. Microbial Pathogenesis 113: 135-143.
- El-Katatny M, Gudelj M, Robra KH, Elnaghy M, Gübitz G (2001) Characterization of a chitinase and an endo-β-1, 3-glucanase from Trichoderma harzianum Rifai T24 involved in control of the phytopathogen Sclerotium rolfsii. Applied Microbiology and Biotechnology 56(1): 137-143.
- Mazrou YS, Makhlouf AH, Elseehy MM, Awad MF, Hassan MM (2020) Antagonistic activity and molecular characterization of biological control agent Trichoderma harzianum from Saudi Arabia. Egyptian Journal of Biological Pest Control 30(1): 4.
- Chet I, Harman GE, Baker R (1981) Trichoderma hamatum: Its hyphal interactions with Rhizoctonia solani and Pythium Microbial Ecology 7(1): 29-38.
- Elad Y, Chet I, Boyle P, Henis Y (1983) Parasitism of Trichoderma on Rhizoctonia solani and Sclerotium rolfsii-scanning electron microscopy and fluorescence microscopy. Phytopathology 73(1): 85-88.
- Gomes EV, Costa MD, De Paula RG, Ricci De AR, Da Silva FL, et al. (2015) The Cerato-Platanin protein Epl-1 from Trichoderma harzianum is involved in mycoparasitism, plant resistance induction and self cell wall protection. Scientific Reports 5(1): 17998.
- De Paula RG, Antoniêto AC, Ribeiro LF, Carraro CB, Nogueira KM, et al. (2018) New genomic approaches to enhance biomass degradation by the industrial fungus Trichoderma reesei. International Journal of Genomics 2018: 1974151.
- Braun H, Woitsch L, Hetzer B, Geisen R, Zange B, et al. (2018) Trichoderma harzianum: Inhibition of mycotoxin producing fungi and toxin biosynthesis. International Journal of Food Microbiology 280: 10-16.
- Chen JL, Sun SZ, Miao CP, Wu K, Chen YW, et al. (2016) Endophytic Trichoderma gamsii YIM PH30019: A promising biocontrol agent with hyperosmolar, mycoparasitism, and antagonistic activities of induced volatile organic compounds on root-rot pathogenic fungi of Panax notoginseng. Journal of Ginseng Research 40(4): 315-324.
- Muniroh MS, Nusaibah SA, Vadamalai G, Siddique Y (2019) Proficiency of biocontrol agents as plant growth promoters and hydrolytic enzyme producers in Ganoderma boninense infected oil palm seedlings. Current Plant Biology 20: 100116.
- Ashwini N, Srividya S (2014) Potentiality of Bacillus subtilis as biocontrol agent for management of anthracnose disease of chilli caused by Colletotrichum gloeosporioides 3 Biotech 4(2): 127-136.
- Thrane C, Tronsmo A, Jensen DF (1997) Endo-1, 3-β-glucanase and cellulase from Trichoderma harzianum: Purification and partial characterization, induction of and biological activity against plant pathogenic Pythium spp. European Journal of Plant Pathology 103(4): 331-344.
- Datsomor O, Yan Q, Opoku-Mensah L, Zhao G, Miao L (2022) Effect of different inducer sources on cellulase enzyme production by white-rot basidiomycetes Pleurotus ostreatus and Phanerochaete chrysosporium under submerged fermentation. Fermentation 8(10): 561.
- Naher L, Fatin SN, Sheikh MA, Azeez LA, Siddiquee S, et al. (2021) Cellulase enzyme production from filamentous fungi Trichoderma reesei and Aspergillus awamori in submerged fermentation with rice straw. Journal of Fungi 7(10): 868.
- Ravindran R, Jaiswal AK (2016) Microbial enzyme production using lignocellulosic food industry wastes as feedstock: A review. Bioengineering 3(4): 30.
- Sulyman AO, Igunnu A, Malomo SO (2020) Isolation, purification and characterization of cellulase produced by Aspergillus niger cultured on Arachis hypogaea Heliyon 6(12): e05668.
- Ridout CJ, Coley-Smith JR, Lynch JM (1986) Enzyme activity and electrophoretic profile of extracellular protein induced in Trichoderma by cell walls of Rhizoctonia solani. Microbiology 132(8): 2345-2352.
- Martin K, McDougall BM, McIlroy S, Chen J, et al. (2007) Biochemistry and molecular biology of exocellular fungal β-(1, 3)-and β-(1, 6)-glucanases. FEMS Microbiology Reviews 31(2): 168-192.
- Zhao J, Xue QH, Niu GG, Xue L, Shen GH, et al. (2013) Extracellular enzyme production and fungal mycelia degradation of antagonistic Streptomyces induced by fungal mycelia preparation of cucurbit plant pathogens. Annals of Microbiology 63(2): 809-812.
- Kim JY, Kwon HW, Yun YH, Kim SH (2016) Identification and characterization of Trichoderma species damaging shiitake mushroom bed-logs infested by Camptomyia pest. Journal of Microbiology and Biotechnology 26(5): 909-917.
- Prasannath K (2017) Plant defense-related enzymes against pathogens: A review. AGRIEAST: Journal of Agricultural Sciences 11(1): 38-48.
- Mahat NA, Muktar NK, Ismail R, Abdul Razak FI, Abdul Wahab R, et al. (2018) Toxic metals in Perna viridis mussel and surface seawater in Pasir Gudang coastal area, Malaysia, and its health implications. Enviro
© 2026 Muhammad Muhammad. This is an open access article distributed under the terms of the Creative Commons Attribution License , which permits unrestricted use, distribution, and build upon your work non-commercially.
Editor In Chief







.jpg)

























Signup for Newsletter

Quick Links
 Editorial Board Registrations
Editorial Board Registrations Submit your Article
Submit your Article Refer a Friend
Refer a Friend Advertise With Us
Advertise With UsOur Recent Edition

.jpg)



Top Editors




.jpg)

















.bmp)
.jpg)
.png)
.jpg)













.jpg)






.png)

.png)




.png)




Financial Support

Sponsors

Latest e-Books

Latest Video

 a Creative Commons Attribution 4.0 International License. Based on a work at www.crimsonpublishers.com.
Best viewed in
a Creative Commons Attribution 4.0 International License. Based on a work at www.crimsonpublishers.com.
Best viewed in