
- About Us
- Guidelines
-

The Author ensures that the research has been conducted responsibly and ethically with adherence to all relevant regulations. read more..
- For Authors
- For Reviewer
- Manuscript Guidelines
- Membership
-
- Journals
- Reprints
- e-Books
- Videos
- Indexing
- Contact Us
 COVID-19
COVID-19
- Submissions



Full Text
Environmental Analysis & Ecology Studies
The Interspecific Interaction Mechanism of Plant Diversity Affecting Alien Species Invasion
Song Gao, Xiao Yan Wang and Jiang Wang*
College of Life Sciences, Taizhou University, China
*Corresponding author: Jiang Wang, Zhejiang Provincial Key Laboratory of Plant Evolutionary Ecology and Conservation, College of Life Sciences, Taizhou University, Taizhou, China
Submission: February 23, 2021; Published: December 1, 2021

ISSN 2578-0336 Volume9 Issue2
Abstract
Interspecies interaction is the basis link among different plants. However, plant interaction has not been pay more attention in many studies of plant diversity-invasibility relationship. In this study, artificial plant communities with different diversity gradients (1, 2, 4 and 8) were established. Two comparative experimental blocks were established with changing the distribution of invasive plants and native plants, i.e., the staggered and non-staggered distribution. The results showed that species richness and functional group richness were significantly negatively correlated with the biomass of S. Canadensis under staggered distribution, while there was no significant relationships under non-staggered distribution. The interspecific competition intensity of S. Canadensis under the staggered distribution was significantly stronger than that under the non-staggered distribution. The results showed that the distribution pattern of native and invasive plants may be an important factor affecting the resistance of plant diversity to invasion of alien plants.
Keywords: Species richness; Functional groups; Solidago canadensis; Invasion; Interspecific interaction
Introduction
Due to serious threat of biological invasion to ecosystem functioning and human health
[1,2], Community invasibility has received more and more attentions [3]. Competition is
considered to be the main interspecific interaction between native and invasive plants [4].
Effect of plant diversity on invader through interspecific interaction between native and
invasive plants at neighborhood scale [5-7]. Therefore, the small-scale spatial structure of
species will certainly affect interspecific interaction. Bufford et al. [8] found that the resource
competition of native plants around invasive plants had an important impact on the growth
and reproduction of invader in the Hawaiian lowland community. Stoll & Prati [9] found that
the aggregation of of same species can reduce the pressure of interspecific competition, and
lead to the survival of less competitive species. Therefore, the different distribution patterns
of plant species may change the impact of native plant communities on invasive plants.
In this study, we construct native plant communities with different species richness and
control competition intensity between native plants and invasive plants by change species
distribution patterns.
We address two questions:
A. Does change of species distribution pattern change intensity of interspecific competition
between native plants and invasive plants?
B. Whether change of interspecific competition intensity change the richness-invasibility
relationship?
Materials and Methods
Eight native species grow in area around Taizhou city (Plantago asiatica L., Solanum nigrum L., Cirsium setosum (Willd.) MB., Kalimeris indica L., Lysimachia fortunei Maxim., Inula japonica Thunb., Polygonum caespitosum BL. and Viola japonica Thunb) were selected, and Solidago canadensis L. were selected as invader. Pots (40cm diameter and 28cm height) with different species richness (1, 2, 4 and 8) were established by randomly selected from eight native species. Sixteen native plants were set in each plot, and the number of each species was divided equally according to the species richness to ensure the cross distribution of different species. Weeding and watering were carried out regularly to maintain original composition. According to the above experimental design, two blocks (control and invasion) were established. In the invasion block, the staggered (Figure 1B) and non-staggered (Figure 1A) distribution treatments of S. canadensis (four plants) were constructed. In September 2017, plant biomass was harvested, separated by species, dried at 80 ℃ for 48h, and weighed. During the experiment, plant height and coverage of each plant in each pots were measured four times.
Figure 1: Distribution of native and invasive plants.
A: Staggered distribution
B: Non-staggered distribution

Data analysis
Growth index of S. canadens is was reflected by the product of plant height and coverage. Interspecific competition index (ICI) was calculated by the following formula:
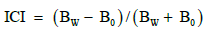
B0: plant biomass in monoculture; BW: plant biomass in mixture.
The average value of interspecific competition index of all
species in community represents interspecific competition
intensity of the community.
Linear regression was used to analyze the effects of species
richness on the biomass and growth potential of S. canadensis.
Paired sample t-test was used to analyze the difference of
interspecific competition intensity between staggered and nonstaggered
distribution treatments. Spss20.0 software was used for
data analysis.
Result
Biomass of invader was negatively correlated to the species richness in staggered treatment (Figure 1) (r = -0.399, n = 51, P = 0.005) but had no significant relationship (r = - 0.153, n = 35, P = 0.358) (Figure 2).
Figure 2: Effects of species richness on biomass of invader under different distributions.

Growth index of S. Canadensis under staggered distribution was significantly lower than that in non staggered distribution (Figure 3, in all four times P <0.05). Moreover, at the third time (r = -0.280, n = 51, P = 0.047) and fourth time (r = -0.376, n = 51, P = 0.007), species richness was negatively correlated to the growth index of S. Canadensis under stagger distribution.
Figure 3: Growth index of S. Canadensis under different distributions.

There was no significant difference in the competition coefficient between staggered and non-staggered communities in the control group (t = 0.979, n = 51, P = 0.332). However, in the invasion group, the competition coefficient of staggered distribution was significantly lower than that of non-staggered distribution (Figure 4) (t = -5.051, n = 51, P < 0.001). There was no significant difference in interspecific competition coefficient between species richness and functional groups (P > 0.05).
Figure 4: Interspecies competition index under different distributions.

Discussion
Results showed that distribution pattern of S. canadensis
changed the species richness-invasibility relationship, which
indicate that species distribution pattern may be a potential
factor that affects species richness-invasibility relationship.
Consistent with the results of Stoll & Prati [9], interspecific
competition intensity between S. canadensis and native plants
under staggered distribution was significantly lower than that
under non-staggered distribution. Effect of plant diversity express
through the interspecific interaction [6,7]. When S. canadensis was
staggered distribution, competition effect of plant diversity cannot
be expressed through neighbor interspecific interactions, and lead
to no significant relationship. Consistent with the result of Bufford
et al. [8], the growth inhibition of native plants on S. canadensis was
stronger under staggered distribution but had no significant effect
under non-staggered distribution.
Moreover, we also found, under the non-staggered distribution,
the inhibition of species diversity on invasion was more obvious
with the extension of time. Therefore, we speculate that the
inconsistent species richness-invasibility relationships of prior
studies may be due to the different distribution patterns of invader,
which affect competition intensity of native plants. This study
provides an theoretical explanation for the debate of species
richness-invasibility relationship.
Acknowledgement
This study was supported by the National Natural Science Foundation of China (31870504), Taizhou University National Funds for Distinguished Young Scientists (2017JQ005, 2019JQ005), the Natural Science Foundation of Zhejiang Province, China (Y13C030003), Basic Public Welfare Research Projects in Zhejiang Province (LGN19C150004).
References
- Funk JL, Cleland EE, Suding KN, Zavaleta ES (2008) Restoration through reassembly: Plant traits and invasion resistance. Trends in Ecology and Evolution 23(12): 695-703.
- Strayer DL (2012) Eight questions about invasions and ecosystem functioning. Ecology Letters 15(10): 1199-1210.
- Koerner SE, Avolio ML, Chang CC, Gary J, Hoover DL, et al. (2015) Invasibility of a mesic grassland depends on the time-scale of fluctuating resources. Journal of Ecology 103(6): 1538-1546.
- Bulleri F, Bruno JF, Benedetti CL (2008) Beyond competition: Incorporating positive interactions between species to predict ecosystem invisibility. Plos biology 6(6): 1136-1141.
- Elton CS (1958) The ecology of invasions. Methuen, London.
- Mack RN, Harper JL (1977) Interference in dune annuals: Spatial pattern and neighbourhood effects. Journal of Ecology 65(2): 345-363.
- Silander JA, Pacala SW (1985) Neighborhood predictors of plant performance. Oecologia 66(2): 256 -263.
- Bufford JL, Lurie MH, Daehler CC (2016) Biotic resistance to tropical ornamental invasion. Journal of Ecology 104(2): 518-530.
- Stoll P, Prati D (2001) Intraspecific aggregation alters competitive interactions in experimental plant communities. Ecology 82(2): 319-327.
© 2021 © Jiang Wang. This is an open access article distributed under the terms of the Creative Commons Attribution License , which permits unrestricted use, distribution, and build upon your work non-commercially.
Editor In Chief







.jpg)
























Signup for Newsletter

Quick Links
 Editorial Board Registrations
Editorial Board Registrations Submit your Article
Submit your Article Refer a Friend
Refer a Friend Advertise With Us
Advertise With UsOur Recent Edition

.jpg)



Top Editors




.jpg)















.bmp)
.jpg)
.png)
.jpg)

















.png)

.png)




.png)




Financial Support

Sponsors

Latest e-Books

Latest Video

 a Creative Commons Attribution 4.0 International License. Based on a work at www.crimsonpublishers.com.
Best viewed in
a Creative Commons Attribution 4.0 International License. Based on a work at www.crimsonpublishers.com.
Best viewed in